
Frozen in Time
Chapter 14. Mammoths Thrive Early in the Post-Flood Ice Age
(매머드는 대홍수 후 빙하기 초기에 번성했다)
출처: https://answersingenesis.org/extinct-animals/ice-age/mammoths-thrive-early-in-the-post-flood-ice-age/
우리는 창세기 대홍수에 의한 기후 변화에 근거하여 대홍수 이후 빙하기 모델을 제안했다. 이제 우리는 털북숭이 매머드의 신비를 탐구 할 수 있는 입장에 서 있다.
털북숭이 매머드의 삶과 죽음을 둘러싼 질문들에 대해 진화론자들과 창조론자들 모두 많은 가설들을 제안했다. 털북숭이 매머드에 관한 정보가 혼란스럽고 대부분의 연구가 러시아어로 출판되어 있기 때문에 창세기 대홍수 모델의 지지자들은 여러 가지 경쟁적인 가설을 세웠다. 일부 창조론자들은 공룡이 사라진 것과 같은 방식으로 대홍수에 의해 매머드가 멸종했다고 설명한다. 그들은 보통 털북숭이 매머드의 멸종과 보존에 대한 메커니즘으로 최소한 시베리아의 영구 동토층에서의 급속한 냉동을 주장한다. 다른 창조론자들은 동일과정설 과학자들도 믿고 있는 것처럼 매머드는 빙하기 동안에 살았고 빙하기가 끝날 때 멸종되었다고 결론을 내렸다. 빙하기가 끝나고 털북숭이 매머드가 멸종되었다고 믿는 사람들 사이에도 급속 냉동 여부를 놓고 의견이 분분하다. 급속 냉동 가설은 다음 장에서 다룰 것이다.
의심의 여지없이, 홍수 이전에는 털북숭이 매머드 뿐 아니라 다른 많은 종류의 코끼리가 있었다. 우리는 매머드를 포함한 이러한 유형의 코끼리 중 일부가 대홍수 동안 화석화되었을 것으로 기대해야 한다. 따라서 매머드는 대홍수 동안에 또는 대홍수 이후에 또는 둘 다에 화석화되었을 가능성이 있다. 이 책의 초점인 털북숭이 매머드는 거의 모두 결빙되지 않은 또는 표면이 얼어붙은 퇴적물에서 발견되며, 이는 그들이 대홍수 이후의 빙하기 동안에 살았다는 것을 나타낸다(부록 4 참조). 이것은 시베리아, 알래스카 및 유콘에 있는 얼어붙은 매머드를 포함한다.
Why were the lowlands of Siberia, Alaska, and the Yukon unglaciated?
(왜 시베리아, 알래스카 및 유콘의 저지대들은 빙하가 없었을까?)
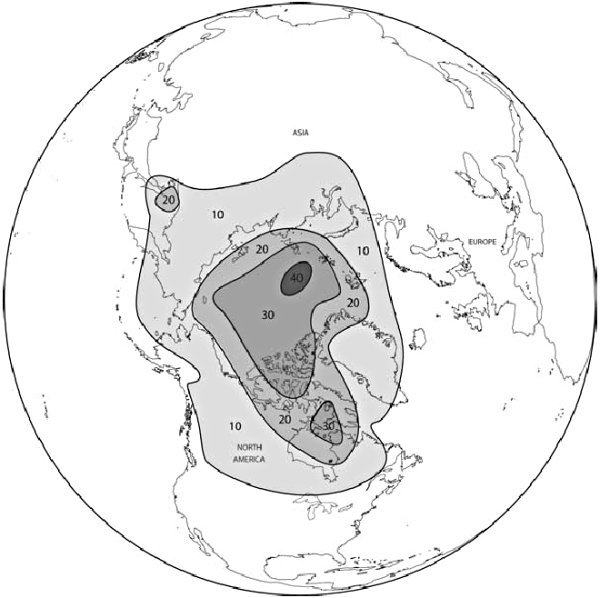
그림 14.1. 100분도로 표시한 극위도 지역에서의 겨울 기온 변화(북극해의 온도를 빙점으로 유지하고 빙모를 제거했다)(Winter temperature changes in centigrade at polar latitudes with just the removal of the Arctic sea ice cap, leaving the temperature of the ocean at freezing.)
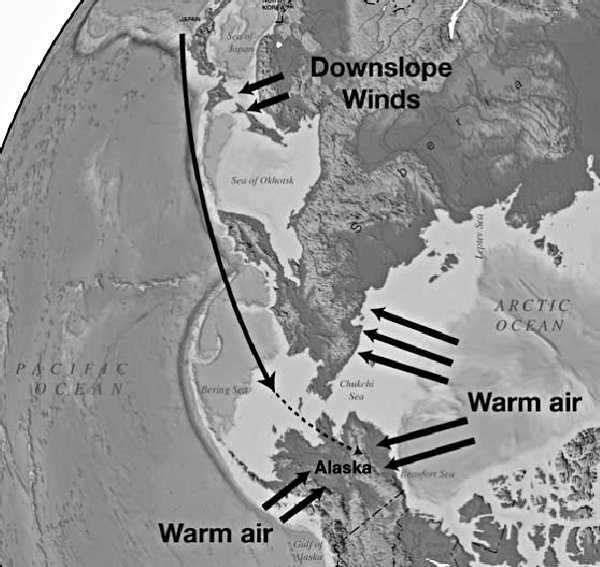
그림 14.2. 주 폭풍의 진로는 동아시아에서 멀어지는 방향이고, 따뜻한 북극해와 북태평양에 의해 따뜻해진 공기가 바다에서 육지로 이동했을 것이다.
시베리아, 알래스카, 그리고 유콘에서 수백만 마리의 털북숭이 매머드가 살았기 때문에, 그들이 빙하기 동안에 어떻게 살 수 있었을까 하는 문제가 자연스럽게 등장한다. 실제로 그들은 빙하가 없는 지역에 살았었는데, 이것은 빙하기에 대한 동일과정설적 관점의 주요 미스테리이다(그림 2.3을 보라). 동일과정설적인 가정을 사용하는 기후 시뮬레이션은 종종 이 지역들이 처음에 빙하가 형성되는 지역으로 나타난다.
이 책에서 제시된 대홍수 이후의 빙하기 모델에서는, 따뜻한 북극해와 태평양 때문에 그 저지대는 빙하가 형성 되지 않았을 것이다. 북극해는 극 위도에 위치하기 때문에 대기에서 열과 습기를 빠르게 잃을 것이고, 겨울에 주변 지역을 따뜻하게 하는 데 도움이 될 것이다.
Newson<1>은 대기 순환 시뮬레이션을 통해 따뜻한 북극해의 극적인 기후 결과를 시연했다. 그는 기후 모델을 사용하여 고위도에서 해빙(Sea Ice)을 제거하는 효과를 연구했는데, 이때 북극해의 표면 온도를 바닷물의 빙점으로 유지했다. 그는 기후 모델의 다른 모든 변수를 동일하게 남겨두고 해빙이 있는 경우와 해빙이 없는 경우의 기후 모델을 실행했다. 두 모델의 실행결과를 비교함으로써 그는 북극해 상부의 겨울 대기 온도가 20°C에서 40°C로(40°F에서 70°F로) 상승한 것을 발견했다! 게다가 더 따뜻한 기온은 인접한 땅으로 퍼져나갔다(그림 14.1). 시베리아와 캐나다의 겨울 기온은 12°C에서 28°C로(20°F에서 50°F로) 상승했다.
대홍수 이후의 빙하기 모델에서와 같이 북극해의 온도가 해수면 빙점보다 훨씬 더 따뜻했다면, 공기의 온도는 Newson의 시뮬레이션보다 훨씬 더 따뜻했을 것이다. 그림 14.2는 북극해와 북태평양 바다의 온난한 물에 의해 가열된 상대적으로 따뜻한 공기의 육지쪽 흐름에 의해 시베리아, 알래스카 및 유콘이 어떻게 따뜻해졌는지를 보여준다. 따뜻한 공기가 시베리아, 알래스카 및 유콘의 저지대에서 빙하가 발달하지 못하게 하고 반면에 그 지역의 산들에는 급속히 빨리 빙하가 형성되게 했을 것이다.
Mammoths spread over Northern Hemisphere early in the Ice Age
(매머드는 빙하기 초기에 북반구에 퍼졌다)
매머드가 시베리아로 퍼지는 것을 막을 얼음이 없었으므로, 오늘날 우리가 발견한 북반구의 나머지 지역으로 어떻게 퍼졌을까?
털북숭이 매머드는 창세기에 기록된 한 종류(Kind)에서 유래한 것으로 보인다(창조생물학적 생물분류체계/baraminology의 하위 분야에서의 매머드를 포함하는 코끼리 종류에 대한 설명은 부록 3을 보라). 두 마리의 털북숭이 매머드가 방주에서 내렸든지, 아니면 털북숭이 매머드가 큰 유전자 풀을 가진 두마리의 코끼리들의 유전자에서 발생했든지, 대홍수 이후 처음에는 천천히 번식했을 것이다. 빙하기가 진행됨에 따라 그들의 숫자가 기하급수적으로 급격히 증가했을 것이다. 700년의 빙하기 동안 개체수가 수천만까지 증가 할 수 있는 충분한 시간이 있었다(이 장의 뒷부분에 나오는 “매머드 개체수 폭발”을 보라). 아라랏산들로부터 유럽과 서아시아로 이주하는 것은 아무런 문제가 되지 않았을 것이다. 매머드가 미국으로 이주하는 것이 가장 큰 도전이었다.
일단 그들이 시베리아에 도착하자 마침내 그들은 알래스카에 퍼질 수 있었다. 베링해와 베링해협 뿐 아니라 시베리아와 알래스카 주변의 대륙붕은 매우 얕다. 육지에서 빙하가 형성되면서 시베리아와 알래스카 사이의 대륙붕이 육지 교량을 제공할 정도로 충분히 해수면이 낮아졌다. 매머드와 다른 많은 동물들이 시베리아에서 베링 육지 교량을 건너서 알래스카와 유콘으로 퍼졌다.<2> 뾰족뒤쥐(shrew)와 들쥐(meadow mouse)와 같이 작은 포유동물들도 그들과 함께 했다. 매우 추운 조건을 견딜 수 없는 동물들은 겨울 기온이 오늘날보다 훨씬 더 따뜻했던 빙하기 초기에 이 경로를 통해 북아메리카로 갈 수 있었다.
유콘 지역으로부터 얼음이 없는 통로를 통해 로키 산맥의 동쪽 경사면을 따라 남동쪽으로 이동했다. 몬태나 주에 도착하자마자 동물들은 자유롭게 번식하여 미국 전역으로 퍼져 나갔고 남쪽을 향했다. 그림 1.6은 빙하기 동안에 북반구에 살았던 털북숭이 매머드의 분포를 보여준다. 컬럼비아 매머드는 일반적으로 털북숭이 매머드 분포 지역의 바로 남쪽 지역부터 중앙아메리카의 코스타리카에까지 분포했다.<3>
이 이동과 확산의 시기 동안, 전체 북반구 기후의 특징은 온화한 겨울과 차가운 여름이다. 그림 14.3은 대홍수 이후 시베리아에 대한 겨울과 여름 기온의 예상 그래프다. 계절간 기온의 변화가 줄어들어 추운 기후를 선호하는 동물 및 식물은 온화한 기후를 선호하는 동물 및 식물과 혼합되었다. 이 부조화의 조합은 빙하기 퇴적물에서 자주 발견되며 빙하기의 또 다른 수수께끼를 설명한다(1장을 보라).
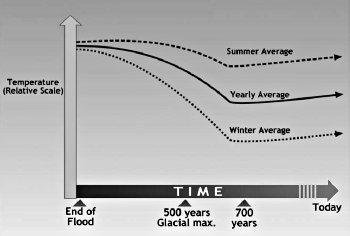
그림 14.3. 시간에 따른 시베리아의 기온 변화 예상 그래프.(Drawn by Dan Lietha of AiG.)
The Ice Age environment of Siberia
(시베리아의 빙하기 환경)
2장에서 우리는 시베리아의 현재 환경이 털북숭이 매머드와 다른 포유류에게 매우 지내기 힘들다는 것을 확실히 설명했다. 매머드가 시베리아와 그 너머로 퍼지도록 하기 위해 대홍수 이후의 빙하기는 어떻게 달랐을까?
방주를 나온 이후, 매머드는 서서히 북동쪽으로 스쳐지나 결국 시베리아에 도달했다. 그들이 도착했을 때, 빙하기가 막 시작했을 것이다. 시베리아 겨울은 온화했으며 여름은 선선했고, 계절간의 차이가 거의 없었다. 환경은 오늘날과 많이 달라서 털북숭이 매머드가 시베리아를 통해 이동할 수 있었다.
겨울이 온난했을 것이라는 추정은 시베리아에서 발견된 사체와 풍부한 뼈(2장을 보라) 뿐만 아니라 다른 데이터에서도 뒷받침된다. 그 증거는 과학자들이 단순히 따뜻함의 증거를 “간빙기”와 “아간빙기(빙하기 중 온난한 기간)” 기간으로 간주하려는 경향에 의해 다소 모호해졌다.<4> Sher<5>는 따뜻한 기후의 지표들을 단순하게 간빙기로 설명하는 것에서 순환논리의 한 요소가 발생한다고 기술한다:
이것은 순환 논리의 위험을 증가시킨다. 예를 들어, 마지막 간빙기가 그 지역에서 가장 따뜻한 기후였다는 결론의 근거가, 이 시기에 퇴적물이 할당된 장소가 인근 지역에서 가장 따뜻한 기후를 나타내는 것으로 보인다는 것을 근거로 결론을 내리는 것은 옳지 않다.
따뜻한 겨울의 징후로는 다음과 같은 것들이 있다: (1)최북단의 영구 동토층 내의 생장 위치에서 큰 나무들이 발견된다.<6> (2)남쪽으로 600 마일(1,000km) 이상 넓은 지역에 자라는 동물의 사체와 관련된 식물들.<7> (3)상당히 따뜻한 기후에 사는 곤충들.<8> (4)오늘날 살아남았지만 훨씬 더 남쪽에서 사는 시베리아와 알래스카의 많은 동물들.<9> (5)북부 시베리아와 알래스카에서 매머드 대초원지대 밑에, 오늘날보다 7~14°F(4~8°C) 더 따뜻한 수온을 나타내는 보다 따뜻한 동물군을 포함하는 해양 퇴적물.<10> 베레소프카(Beresovka) 매머드의 위장에 있는 식물은 처음에는 현재의 툰드라 기후를 지지한다고 주장되었지만, 재분석으로 결과 그것은 훨씬 더 남쪽에서 자랐던 것이다.<11> 미나리 아재비와 같이 오늘날 시베리아에서 자라는 식물도 더 따뜻한 기후에서도 자라는 것이다.
시베리아의 동물과 환경은 빙하기 초기에 온난한 겨울이 있었다는 것을 나타낸다(그림 14.3). 북극해와 북태평양 바다의 습한 공기가 육지 방향으로 이동했기 때문에 대홍수 직후 습한 기후가 발생했을 것이다(그림 14.2를 보라). 따뜻한 공기가 더 많은 수증기를 보유하고 따뜻한 북극해에서 많은 증발이 발생했기 때문에 대홍수 이후 시베리아, 알래스카 및 유콘지역에 강수량이 많았을 것이다. 큰 강수량은 빙하기 초기에 이 지역들에 울창한 숲을 만들었을 것이고, 이것은 지표 퇴적물 내에서 나무와 그루터기가 한꺼번에 발견되면서 명백히 입증된다.
빙하기가 진행됨에 따라 북극해는 점차 냉각되어 증발량이 현저히 줄어들었다. 또한 겨울과 여름 온도가 추워짐에 따라 서서히 대기가 건조해졌다. 화산 먼지와 가스로 인해 여름이 평상시보다 더 시원했을 것이기 때문에 여름철 소나기와 뇌우가 대부분 억제되었을 것이다. 대규모의 시원한 계절 폭풍은 여름에 약해졌고, 여름 소나기가 오늘날보다 적었기 때문에 빙하기 중기 및 후기 동안의 여름은 일반적으로 건조했을 것이다. 따라서 시베리아의 기후는 빙하기가 시작된 이래 점차적으로 건조해졌을 것이다(그림 14.4). 환경은 일반적으로 숲이 우거진 풍경에서 빙하기의 중반 경에 광대한 초원 또는 대초원 환경으로 바뀌었을 것이다. 초원은 오늘날 북미의 고원 지대에서와 같이 상대적으로 온화한 겨울 기온, 풍부하고 시원한 계절적 강수량 및 건조한 여름 기후 속에 생겨난다. 대초원 환경에서의 풀은 오늘날 습지에서 자라는 식물과 달리 포유류가 먹을 수 있는 초목이었다. 포유동물은 시베리아의 넓은 영역에서 많은 풀을 먹었을 것이다. 이 시나리오는 동일과정설 과학자들이 “생산성 역설(productivity paradox)”이라고 부르는 것을 해결할 수 있다. 이것은 이용할 수 있다고 생각되는 음식의 양에 비해 너무 많은 동물들이 번성하는 상태이다.
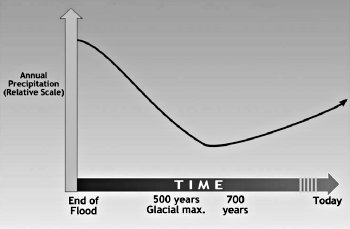
그림 14.4. 시간에 따른 시베리아 연간 강수량 추정 그래프 (Drawn by Dan Lietha of AiG.)
심지어 북극해 표면이 해수 빙점까지 온도가 떨어진 후에도 여전히 빙결되지 않았을 것이다. (빙점에서 Newson의 시뮬레이션이 가장 적합할 것이다.) 이에 대한 이유는 여러 가지가 있지만, 이는 주로 바닷물 결빙이 어렵다는 것과 북극해의 물 순환으로 인한 것이다. 북극해는 빙하기 내내 그리고 해빙기(해빙수가 소금물 위로 떠다니고 얼어붙을 때)까지 얼음이 없는 상태로 남았을 가능성이 크다.
No permafrost at the beginning(초기에는 영구 동토층이 없었다)
오늘날 시베리아의 영구 동토층은 여름에 늪을 만들고 포유류가 시베리아로 이주하는 것을 막아준다. 빙하기 동안 시베리아에 많은 포유동물이 서식할 수 있도록 빙하기의 겨울은 영구 동토층의 성장을 막을 만큼 충분히 따뜻했을 것이다.
오늘날 영구 동토층은 불연속적 구역과 연속적인 구역으로 나뉘어져 있다. 불연속적인 영구 동토층은 연간 평균 기온이 약 30°F(-1°C)일 때 형성되며, 연속적인 영구 동토층은 연평균 기온이 20°F(-7°C) 혹은 그 이하로 떨어질 때 형성된다.<12> 시베리아의 빙하가 없는 영역의 연속적인 영구 동토층은 아마도 빙하기 후반까지 발생하지 않았을 것이다(그림 14.5).
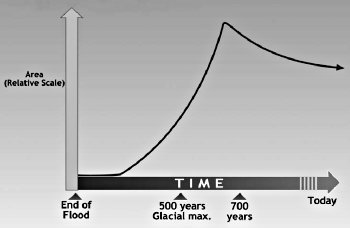
그림 14.5. 시간에 따른 시베리아 영구 동토층 추정 면적(Drawn by Dan Lietha of AiG.)
빙하기 초기와 중기 동안 거의 혹은 전혀 영구동토층이 형성되지 않았을 것이라는 추정은 동일과정설 과학자들에게는 생각할 수 없는 것이다. 그러나 실제로 이것이 사실이라는 상당한 증거가 있다. 특별히 털북숭이 매머드와 사이가산양(saiga antelope) 등과 같이 시베리아에 살았던 많은 포유동물들은 늪지대에서 걷는 데 큰 어려움을 겪었을 것이다(2장을 보라).
작은 발굽을 가진 사이가산양(saiga antelope)은 영구 동토층이 없다는 강력한 증거이다. 영양은 지금 남부 시베리아의 대초원에 산다. 그들의 고도로 전문화된 발굽은 개방적이고 평평한 지형을 선호하며 늪지대나 많은 눈이 쌓인 곳을 여행 할 수 없다.<13> Sher<14>는 다음과 같이 말한다: “홍적세의 사이가산양(saiga antelope)은 오늘날처럼 흐물흐물한 땅, 늪지대, 깊은 눈 위로 이동할 수 없다.” 그리고 빙하기 사이가산양(saiga antelope)의 화석은 시베리아 전역, 심지어 노보시비르스크 제도(New Siberian Islands), 알래스카 및 유콘에서도 발견된다(그림 2.4를 보라). 알래스카에서는 내륙과 북극해 근처에서 발견된다. 유콘에서 유일한 발견은 맥켄지 삼각주(MacKenzie delta)의 동쪽에 위치한 북극 해안 근처이다. 이것은 북극 해안을 따라 북서쪽 로렌타이드 빙상(Laurentide ice sheet)의 발달이 지연되었음을 나타낸다. 이 같은 지연은 동일과정설의 빙하기 모델에서 이상한 것처럼 보이지만, 대홍수 이후의 빙하기 모델에서는 북쪽에 따뜻한 북극해가 있기 때문에 예상되는 것이다.
일부 진화론과학자들은 땅이 영구적으로 얼어붙었다고 추론하여 사이가산양 화석을 조화시키려고 시도했다.<15> 이런 식으로 사이가산양은 빙하기에 시베리아 전역으로 퍼질 수 있었다는 것이다. 땅이 영구적으로 얼어붙었다면, 사이가산양과 다른 동물들은 무엇을 먹었을까? 동물들이 추운 기후에서도 살아남을 수 있는지 여부는 의심스럽다. 빙하기에 먼 북쪽에 살았던 많은 동물들은 이제 더 온화한 기후를 가진 훨씬 더 남쪽에 살고 있다.
비버 화석과 비버가 자른 나무도 빙하기의 일정 기간 동안 시베리아, 알래스카 및 유콘에서 영구 동토층이 없었다는 것을 나타낸다. (이것은 비버가 자른 나무의 해석이 정확하다고 가정한다.) Quackenbush<16>은 북서 알래스카에서 비버 댐의 얼어붙은 잔해를 발견했다고 믿고 있다. 비버는 영구 동토층을 좋아하지 않는다. 왜냐하면 영구 동토층은 여름에는 상부 2 피트(60cm)만 녹고 겨울에는 다시 얼기 때문이다. 오소리와 흰 족제비와 같은 깊은 굴에 사는 동물은 빙하기에 시베리아에 살았으며 일정 기간 동안 영구 동토층이 없다는 증거를 제공한다.
동물 데이터의 가장 직설적인 해석은 빙하기의 겨울이 훨씬 더 따뜻했으며 영구 동토 층은 빙하기 말까지 생겨나지 않았다는 것이다.
Hippos associated with woolly mammoths and reindeer
(하마는 털북숭이 매머드 및 순록과 관련이 있다)
하마는 북서부 유럽에서 순록과 털북숭이 매머드 화석과 함께 발견된다(1장을 보라). 이것은 아마 부조화의 조합 중 가장 드라마틱한 예이지만, 그러한 부조화의 조합은 빙하기에 일반적인 것이었다. 대홍수 후의 온난한 겨울과 선선한 여름이라는 독특한 기후는 급격하게 빨리 빙하기를 가져왔고, 부조화의 조합을 만들어냈다.
특히, 추운 기후 동물과 관련된 하마의 상황은 북서 유럽이 빙하기 초반 온난한 육지 흐름 가운데 놓여있었음을 깨달음으로써 설명 될 수 있다. 하마는 유럽을 가로질러 남부 잉글랜드로 접어들면서 쾌적한 환경을 발견했을 것이다. 빙하기가 진전되고 해수가 냉각됨에 따라 북서 유럽의 기후는 시간이 지나면서 점차 추워졌을 것이다. 빙하기가 끝날 무렵, 영국 섬의 산과 저지대에 마침내 빙하가 형성되었다. 이 기간 동안, 추위를 선호하는 동물들은 얕은 북쪽 해상의 또 다른 육지교량을 거쳐 잉글랜드 남부로 이주했다. 그곳에서 그들은 그 지역을 점령하고 있으며 기후에 스트레스를 받는 하마들과 섞였다. 이것은 어떻게 냉혹한 추위를 잘 견딜 수 있는 몇몇 동물들이 하마와 임시로 살았고 결국 함께 묻혔는지를 설명해 준다.
부조화의 조합은 여전히 동일과정설적인 과학자들에게는 거의 완벽한 비밀이지만, 대홍수 이후 빙하기 모델에서는 충분히 예상된다. 그러나 동일과정설적인 과학자들에게 가설이 없지는 않다. 부록 2는 빙하기 부조화의 조합에 대한 동일과정설적인 네 가지 주요한 설명을 비평한다.
Mammoth population explosion(매머드 개체수 폭발)
시베리아에만 약 1 천만 마리의 털북숭이 매머드가 묻혀있다. 단지 700 년의 빙하기를 거치면서 그러한 대규모 개체수가 성장하기에 충분한 시간이었는지 사람들은 의문을 제기한다. 우리는 아프리카 코끼리의 번식 습성을 조사함으로써 대홍수 이후의 엄청난 개체수 폭발을 예측할 수 있다.
매머드가 코끼리 종류의 한 구성원일 가능성이 높기 때문에 현대 코끼리는 털북숭이 매머드에 대한 좋은 유사체이다(부록 3을 보라). 매머드는 현대 코끼리, 특히 아시아 코끼리와 비슷한 외형과 크기를 가지고 있다.<17> 그 둘 다 특수한 엄니, 몸통, 기둥 모양의 다리가 비슷하다. 최근 네팔에서 다양한 아시아 코끼리를 관찰했는데, 머리 뒤에서 거대한 혹과 매머드의 전형적인 경사진 등이 관찰되었다.<18> 매머드의 뼈는 현대 코끼리와 비슷한 길이지만 더 거대하다.<19> 매머드 DNA는 코끼리 DNA와 유사하다.<20> 콜로라도 고원의 동굴과 벽감(alcove)에 보존된 매머드 배설물<21>과 시베리아 매머드의 위속 내용물은 코끼리와 매머드가 동일한 후위 소화를 하는 것을 나타낸다.<22> 따라서 매머드는 현대의 코끼리와 유사한 사회적 특징, 생식 전략, 성장 패턴, 먹이 스타일을 가졌을 가능성이 크다.<23>
코끼리의 생식률은 상당히 다양하다.<24> 코끼리는 10~23 세까지 성적 성숙에 이르지 못한다.<25> 그들은 50년에서 60년까지 산다. McDonald<26>는 현대 코끼리는 일반적으로 한 배에 1 마리의 새끼를 낳고 5 년마다 출산하는데 시간은 더 짧을 수 있다고 말한다. Haynes<27>는 짐바브웨의 황게(Hwange) 국립 공원에서 코끼리 암컷 한 마리가 3년에서 9년 동안 새끼를 낳을 수 있으며 평생 5마리에서 15마리의 새끼를 낳을 수 있다고 기록했다. Eltringham<28>은 코끼리가 일반적으로 4-5 년 간격으로 송아지를 생산하며 그때 쌍둥이가 태어날 확률이 1.35%라고 말한다. 그러나 어떤 이들을 코끼리가 2~3 년마다 출산 할 수 있다고 제안하기도 했으며, 한 동물원 코끼리의 경우 출생한지 2년 5개월이 지나 처음으로 출산하기도 했다고 한다.<29> 코끼리가 당장 임신을 하지 않으면 2개월 후에 또 다른 주기가 생긴다. 그래서 암컷은 2년에서 5년을 더 기다릴 필요는 없지만, 임신에 실패한 후 곧 임신이 될 수 있다. 임신기간은 21~22개월이며 새끼는 2년 동안 젖을 빤다. Fisher<30>는 엄니의 성장 속도를 토대로 매스토돈(mastodons)이 10 세에 성적 성숙을 하고 2~6년마다 출산 할 수 있다고 결론지었다.
번식률은 특히 환경이 좋을 때, 개체수가 적을 때 혹은 동물들을 정기적으로 사냥할 때 늘어난다.<31> 성숙한 코끼리에게는 인간을 제외하고는 천적이 없지만<32> 새끼들은 포식자의 대상이 된다. 새끼의 경우 특히 수컷은, 가뭄 기간 동안 사망률이 증가한다.<33> 새끼가 죽으면 어미 코끼리는 예상보다 빨리, 약 20개월 후에 새끼를 밴다.<34> 코끼리는 다양한 환경에 잘 적응하고<35> 아프리카의 건조한 계절 동안 짧은 거리를 이동한다. 그들은 대부분 풀을 뜯어 먹지만 나무와 덤불에서 잎을 찾아 먹을 수도 있다.<36> 그래서 코끼리는 척박한 환경에 매우 강하며 좋은 환경에서 상당히 빨리 번식할 수 있다.
이러한 변수에 기초하여, 빙하기 동안 털북숭이 매머드의 개체수의 증가를 추정하는 것은 어려울 것이다. 해결할 수 있는 지름길은 데이터가 있는 다양한 아프리카 코끼리 무리의 번식률과 비교하여 털북숭이의 매머드 배가율을 추정하는 것이다. Haynes<37>는 황게(Hwange) 국립 공원에서 코끼리 개체수가 20년 만에 13,000에서 22,000으로 증가했다고 보고했다. 이것은 약 25년에 두 배로 증가하는 것이다. 이 번식률은 밀렵과 가뭄에도 불구하고 유지된다. 매머드가 이 비율로 증가한다면 홍수 이후 550년 이내에 살아있는 매머드가 800만 마리가 될 것이다.<38> 그러나 코끼리의 번식률이 현저하게 빠를 수 있다. Mithen<39>은 아프리카 코끼리의 경우 매년 3~5%의 개체 증가가 통상적으로 보고된다고 말한다. 4퍼센트 비율로, 매 18년 마다 두 배씩 증가하면, 500년 안에 매머드가 6억 5천 7백만 마리가 될 것이다. Mithen<40>은 또한 남아프리카의 Addo 국립 공원에서 27년 동안 개체수가 매년 7% 증가했다고 발표했다. 이는 오늘날 아프리카에서 가능한 최대 성장률일 가능성이 높다. 이것은 이전에 매머드에 사용했던 추정치인 약 10 년에 두 배가 되는 시간이다.<41> 이 배가율로 추정하면 대홍수 후 300년 후에 13억 마리의 매머드가 살고 있었을 것이다! 이 추정치는 아마 매머드의 번식율에 가까울 것이며, 특히 포식자의 수가 아직 적고 환경이 더 이상적이었던 대홍수 후 초기 몇 년 동안은 더욱 그러하다. 따라서, 700 년의 빙하기 동안 수백만 마리의 털북숭이 매머드가 이었었다는 것이 문제가 되지 않는다. 그림 14.6은 대홍수가 끝난 후 빙하기가 끝날 때까지의 매머드 수에 대한 타임 라인을 보여준다.
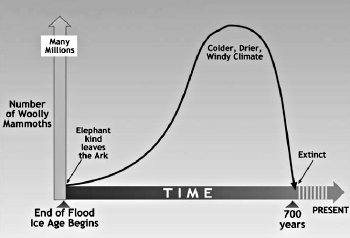
그림14.6. 대홍수부터 현재까지의 시간에 따른 털북숭이 매머드 개체수.(Drawn by Dan Lietha of AiG.)
시베리아와 알래스카에 있는 매머드의 유해들은 포식자의 흔적을 찾을 수 없을 정도로 건강해 보였다. Guthrie<42>는 치아 마모 상태를 토대로 대부분의 매머드가 장수했다고 설명한다. 위의 번식률은 빙하기 말기에 살고 있는 매머드의 잠재적 개체수에 대한 추정치이다. 그 숫자에는 잠재적으로 화석이 될 수 있는 빙하기에 살았던 더 많은 개체들을 포함하지 않는다.
Footnotes(각주)
1. Newson, R.L., Response of a general circulation model of the atmosphere to removal of the Arctic ice-cap, Nature 241:39–40, 1973.
2. Oard, M.J., An Ice Age Caused by the Genesis Flood, Institute for Creation Research, El Cajon, CA, pp. 84–86, 1990.
3. Agenbroad, L.D. and L. Nelson, Mammoths: Ice Age giants, Lerner Publications Company, Minneapolis, MN, p. 22, 2002.
4. For example, in Ukraintseva, V.V., Vegetation of warm Late Pleistocene intervals and the extinction of some large herbivorous mammals, Polar Geography and Geology 4:189–203, 1981.
5. Sher, A.V., Problems of the last interglacial in Arctic Siberia, Quaternary International 10–12:215, 1991.
6. Quackenbush, L.S., Notes on Alaskan mammoth expedition of 1907 and 1908, Bulletin of the American Museum of Natural History 25:126, 1909. Kaplina, T.N., and A.V. Lozhkin, Age and history of accumulation of the “ice complex” of the maritime lowlands of Yakutia; in: Late Quaternary environments of the Soviet Union, A.A. Velichko (Ed.), University of Minnesota Press, Minneapolis, MN, pp. 147–151, 1984.
7. Guthrie, R.D., Frozen fauna of the mammoth steppe — The story of Blue Babe, University of Chicago Press, Chicago, IL, p. 185, 1990. Ukraintseva, Vegetation and extinction. Ukraintseva, V.V., Vegetation cover and environment of the “Mammoth Epoch” in Siberia, Mammoth Site of Hot Springs, South Dakota, Inc., Hot Springs, SD, 1993.
8. Matthews Jr., J.V., Quaternary environments at Cape Deceit (Seward Peninsula, Alaska): Evolution of a tundra ecosystem, Geological Society of America Bulletin 85:1364–1365, 1974. Kiselev, S.V., and V.I. Nazarov, Late Pleistocene insects; in: Late Quaternary environments of the Soviet Union, A.A. Velichko (Ed.), University of Minnesota Press, Minneapolis, MN, pp. 223–226, 1984. Berman, D., S. Armbruster, A. Alfimov, and M. Edwards, Subarctic steppe communities in Beringia; in: Bridges of the science between North America and the Russian Far East, 45th Arctic science conference, section 2 — Beringia revisited: Recent discoveries and interpretations, p. 10, 1994.
9. Guthrie, R.D., Frozen fauna of the mammoth steppe — The story of Blue Babe, University of Chicago Press, Chicago, IL, pp. 310–311, 1990.
10. Larsen, E., S. Funder, and J. Thiede, Late Quaternary history of northern Russia and adjacent shelves — A synopsis, Boreas 28:6, 1999. Mangerud, J., J.I. Svendsen, and V.I. Astakhov, Age and extent of the Barents and Kara ice sheets in Northern Russia, Boreas 28:46–80, 1999.
11. Ukraintseva, V.V., Vegetation cover and environment of the “Mammoth Epoch” in Siberia, Mammoth Site of Hot Springs, South Dakota, Inc., Hot Springs, SD, p. 18, 1993.
12. Washburn, A.L., Geocryology: A survey of periglacial processes and environments, John Wiley & Sons, New York, p. 26, 1980.
13. Sher, A.V., Fossil saiga in northeastern Siberia and Alaska, International Geology Review 10(11):1247–1260, 1968. Harington, C.R., Pleistocene saiga antelopes in North America and their paleoenvironmental implications; in: Quaternary Paleoclimate, W.C. Mahaney (Ed.), Geo Abstracts, Norwich, England, pp. 193–225, 1981.
14. Sher, A.V., Fossil saiga in northeastern Siberia and Alaska, International Geology Review 10(11):1252, 1968.
15. Sher, A.V., Fossil saiga in northeastern Siberia and Alaska, International Geology Review 10(11):1247–1260, 1968.
16. Quackenbush, L.S., Notes on Alaskan mammoth expedition of 1907 and 1908, Bulletin of the American Museum of Natural History 25:87–130, 1909.
17. Haynes, G., Mammoths, mastodonts, and elephants, Cambridge University Press, Cambridge, NY, 1991.
18. Sarfati, J., Mammoth — Riddle of the Ice Age>, Creation 22(2):10–15, 2000.
19. Haynes, Mammoths, mastodonts, and elephants, pp. 22–24.
20. Höss, M., S. Pääbo, and N.K. Vereshchagin, Mammoth DNA sequences, Nature 370:333, 1994.
21. Agenbroad, L.D., and J.I. Mead, Quaternary geochronology and distribution of Mammuthus on the Colorado Plateau, Geology 17:861–864, 1989.
22. Haynes, Mammoths, mastodonts, and elephants, p. 59.
23. Haynes, Mammoths, mastodonts, and elephants, p. 106.
24. Laws, R.M., I.S.C. Parker, and R.C.B. Johnstone, Elephants and their habitats — The ecology of elephants in North Bunyoro, Uganda, Clarendon Press, Oxford, pp. 204–227, 1975.
25. Pilgram, T., and D. Western, Inferring hunting patterns on African elephants from tusks in the international ivory trade, Journal of Applied Ecology 23:503–514, 1986.
26. McDonald, J.N., The reordered North American selection regime and Late Quaternary megafaunal extinctions; in: Quaternary extinctions: A prehistoric revolution, P.S. Martin and R.G. Klein (Eds.), University of Arizona Press, Tucson, AZ, pp. 421, 428, 1984.
27. Haynes, Mammoths, mastodonts, and elephants, p. 65.
28. Eltringham, S.K., Elephants, Blandford Press, Dorset, England, 1982.
29. Ibid., p. 86.
30. Fisher, D.C., Extinction of proboscideans in North America; in: The Proboscidea — Evolution and palaeoecology of elephants and their relatives, J. Shoshani and P. Tassy (Eds.), Oxford University Press, New York, p. 310, 1996.
31. Laws, Parker, and Johnstone, Elephants and their habitats. Eltringham, Elephants, pp. 84–88. Lee, P.C., and C.J. Moss, Early maternal investment in male and female African elephant calves, Behavioral Ecology and Sociobiology 18:358, 1986. Haynes, Mammoths, mastodonts, and elephants, p. 65. Ward, P.D., The call of distant mammoths — Why the Ice Age mammoths disappeared, Springer-Verlag, New York, p. 219, 1997.
32. Ward, P.D., The call of distant mammoths — Why the Ice Age mammoths disappeared, Springer-Verlag, New York, p. 132, 1997.
33. Lee, P.C., and C.J. Moss, Early maternal investment in male and female African elephant calves, Behavioral Ecology and Sociobiology 18:353, 1986.
34. Lee and Moss, Early maternal investment, p. 358. Fisher, Extinction of proboscideans, p. 310.
35. Ward, Call of distant mammoths, p. 133.
36. Eltringham, Elephants, pp. 92–96.
37. Haynes, Mammoths, mastodonts, and elephants.
38. Sarfati, J., How did millions of mammoth fossils form? Creation 21(4):56, 1999.
39. Mithen, S., Simulating mammoth hunting and extinction: Implications for the Late Pleistocene of the central Russian plain; in: Hunting and animal exploitation in the Late Palaeolithic and Mesolithic of Eurasia, G.L. Peterkin, H.M. Bricker, and P. Mellars (Eds.), The American Anthropological Association, USA, p. 170, 1993.
40. Ibid.
41. Oard, Ice Age, p. 83.
42. Guthrie, Frozen fauna, p. 25.
 Frozen in Time(15장) 시베리아 매머드는 급속 냉동 되었을까? - ...
Frozen in Time(15장) 시베리아 매머드는 급속 냉동 되었을까? - ...
 Frozen in Time(13장) 빙하기 동안 인류는 어디에 있었을까? - Mi...
Frozen in Time(13장) 빙하기 동안 인류는 어디에 있었을까? - Mi...