
Frozen in Time
Chapter 2 Why Live in Siberia(왜 시베리아에 살았을까?)
출처: https://answersingenesis.org/extinct-animals/ice-age/why-live-in-siberia/
무엇이 수백만 마리의 털북숭이 매머드를, 겨울에는 일반적으로 몹시 춥고 여름에는 매우 위험한 습지가 되는 그 먼 북쪽으로 몰아갔는가? 그리고 그들이 거기 있는 동안 무엇을 먹었는가?
털북숭이 매머드의 죽음에 관한 이론을 탐구하기 전에 몇 가지 예비 질문에 답할 필요가 있다. 가장 중요한 것은, 무엇이 수백만 마리의 털북숭이 매머드를, 겨울에는 일반적으로 몹시 춥고 여름에는 매우 위험한 습지가 되는 그 먼 북쪽으로 몰아갔는가? 그리고 그들이 거기 있는 동안 무엇을 먹었는가?
The great Siberian death wish(시베리아의 거대한 죽음의 소원)
1970년대에 사향소가 시베리아 북부의 타이미르(Taimyr) 반도와 북극해에 위치한 랭글(Wrangel) 섬에서 다시 발견 되었다.<1> 만약 털북숭이 매머드와 다른 동물들을 클로닝(복제) 같은 것을 통해 되살릴 수 있다면, 그들은 시베리아에서 생존할 수 있을까? 일부 과학자들은 심지어 냉동된 매머드 사체의 세포들을 아시아 코끼리의 암컷에게 주입하여 잡종을 만들어 낼 수 있다고 제안하기도 한다.
시베리아의 겨울은 혹독한 추위로 잘 알려져 있다(아래의 그림 2.1을 보라). 시베리아의 겨울 평균 기온은 화씨 영하 0도 이하(섭씨 영하 18도 이하)이다. 하지만 겨울에 최저 기온은 (섭씨나 화씨 모두) 영하 40도 이하로 내려가는 것이 보통이다. 북반구에서 기록된 최저 기온은 베르코얀스크(Verkhoyansk)에서 화씨 영하 90도(섭씨 영하 68도)이다.<2> 큰 포유류들이 보통 상당한 추위를 견딜 수 있기는 하지만, 그들이 겨울의 강한 눈보라와 극심한 냉기를 견딜 수 있었을까? 시베리아의 이런 추운 기간은 1년에 거의 9개월 동안 지속된다.
흥미롭게도, 오늘날 시베리아에는 큰 포유류가 거의 살고 있지 않다. Vereshchagin과 Baryshnikov<3>는 털북숭이 매머드에 관한 일부 러시아 과학자들의 의견을 이렇게 대변한다:
“짙은 눈보라를 동반한 매서운 바람이 부는 현재 유라시아의 북극 툰드라에는 매머드가 있을 곳이 없을 것이다.”
주류 과학자들은 빙하 시대 동안 겨울 기온이 화씨 영하 10도에서 20도(섭씨 6도에서 12도) 정도 더 내려갔을 가능성이 있다고 믿고 있는데, 이는 생존을 더욱 어렵게 만드는 조건이 된다.
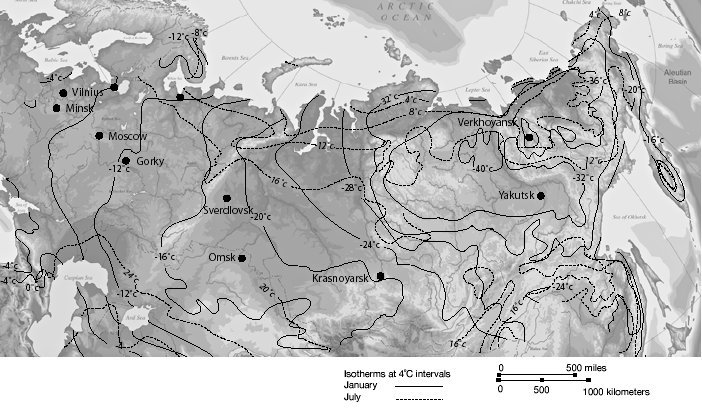
그림 2.1. 1월과 7월의 평균 기온 분포
반면에, 여름은 거의 영구적인 햇빛 때문에 꽤 따뜻해 질 수 있다. Digby<4>는 시베리아의 Yakutsk에서 보낸 일주일을 이렇게 묘사하고 있다. ‘수은주는 결코 화씨 영상 80도 (섭씨 영상 27도) 이하로 떨어지지 않았어요. “밤”에도 낮에도 말입니다! 동물들이 겨울에 살아남았다고 가정하면, 여름은 편안했을 것입니다.’
어떤 사람들은 털북숭이 매머드가 따뜻한 봄과 여름 동안에 시베리아로 쉽게 이주할 수 있다고 생각했다. 초목이 무성한 여름이 지난 후 겨울이 오기 전, 그들이 다시 가을에 남쪽으로 이동했을 것으로 추정 했다. 비록 몇몇 과학자들이 이 가능성에 의미를 둘지라도<5>, 이 이론은 치명적인 결함을 가지고 있다. 매머드는 겨울이 되기 전에 수천 마일을 이주해야만 하므로 이로 인해 매머드는 상당한 에너지를 소모 했을 것이다<6>. 매머드는 다리가 무거워서 다른 포유동물들보다 걷는데 훨씬 더 많은 에너지가 필요하다. Howorth<7>는 시베리아 북동부에 있는 어떤 동물이라도 추운 겨울을 피하기 위해 더 멀리 옮겨야 한다는 점을 지적한다. 그들 경로의 바로 남쪽에는 북태평양이 있어서, 매머드가 남쪽으로 가기 위해서는 서쪽으로 몇 천 마일을 더 멀리 여행해야 했다. 이주 이론의 또 다른 문제점은, 봄에 시베리아 북부로 다시 돌아온다는 것이 이치에 맞지 않다는 것이다. 시베리아 남부나 더 먼 남쪽에서 적절한 식량 공급이 가능했을 것이다<8>. 따라서 북쪽으로 이동할 필요가 없었을 것이다. 마지막 문제점으로 매머드는 임신 기간이 22개월 정도 되기 때문에 이런 종류의 연간 이주는 상당히 위험한 도전이 될 수 있다. 매머드 전문가인 Gary Haynes는 이런 거대한 이주에 대해 다음과 같이 요약했다<9>:
매머드는 순록이 여행하듯이 거의 일년이 걸리는 왕복 여행을 할 수 없었을 것이다. 왜냐하면, 장비목의 동물(역주: 코가 긴 동물)은 식량 공급 요구량이 너무 많아 지속적인 여행을 할 수 없으며, 무리의 더 작은 멤버들, 특히 12세 이하의 젖을 뗀 새끼들의 경우, 나이 든 어른들과 같은 속도로 여행을 할 수 없었을 것이기 때문이다. 게다가, 매머드의 임신 기간이 적어도 22개월이 되었고 임신한 암컷들은 임신 기간 동안 두 번이나 연간 이주를 했어야 했다. 이러한 연간 이주는 그들 자신과 자궁 내 태아에게 견딜 수 없는 부담을 지우게 된다.
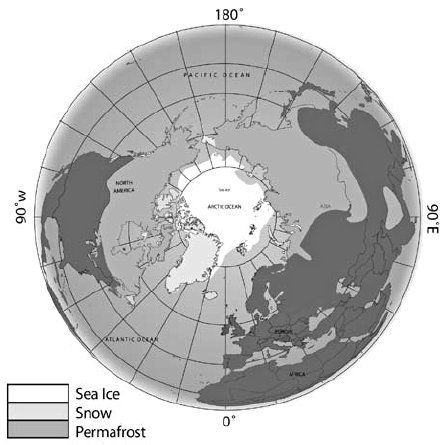
그림 2.2. 북반의 영구 동토층 지도 (영구 동토층은 일반적으로 연평균 기온이 -6ºC ~ -8ºC 인 등온선의 북쪽에 위치한다.)
여름 이주가 가능하지 않을 것이라는 또 다른 압도적인 이유가 있다. 여름 늪지가 있는 기간 동안 여행하는 것은 매머드에게는 치명적이었을 것이다. 시베리아는 현재 수백 피트(100m 이상) 깊이의 흙이 영구히 얼어 있는 영구 동토 지역에 위치하고 있다. 매 여름마다 상부의 1~2피트(약 0.5미터) 정도가 녹는다. 그 아래의 땅은 계속 얼어붙어 있다. 지표층에서 녹은 물은 거대한 습지대와 물이끼가 있는 소택지를 형성한다. 결과적으로 여름 여행은 사람과 짐승 모두에게 힘든 일이 된다<10>. Pfizenmayer<11>는 그의 팀이 여름 동안 베레소프카(Beresovka) 매머드가 있는 곳까지 시베리아를 횡단 여행하는 데 엄청난 노력을 기울였다고 설명한다. 한 겨울 중반이 되어서야 그의 팀은 매머드를 끌어 낼 수 있었다. 그는 끈적이는 습지 진흙이 많은 동물 떼를 죽였을 것이라고 결론 내렸다. Tolmachoff<12>는 이 끈적한 진흙 몇 인치(약 5cm)로 인해 사람이 툰드라를 통행할 수 없었을 것이고, 1피트 혹은 그 이상의 진흙 수렁은 매머드를 가두기까지 했을 것이라고 말한다. 특히 털북숭이 매머드는 기둥 모양의 다리 구조와 뻣뻣한 다리 운동 인해 늪같은 지반을 헤쳐 나가는 데에 어려움을 겪었을 것이다<13>. 또한 최대 보폭 길이를 약간 초과하는 어떠한 도랑도 통과하지 못했을 것이다. Vereshchagin<14>는 분명하게 선언한다: “매머드와 들소는 오늘날 시베리아와 같은 툰드라 지역에서 살 수 없었을 것이다.”
사향 소, 순록, 무스 같은 거대 포유류들은 오늘날 높은 북극 지방에서 살고 생존하고 있다. 하지만 이 초식 동물들은 늪지대에 살기 적합하며, 오늘날 시베리아에서 자라는 식물들을 먹을 수 있다. 순록은 눈 속에서 냄새를 맡아 이끼를 찾아내서 먹는다. 그들은 더 심한 겨울이 오기 전에 남쪽으로 이주한다. 그들의 발굽은 눈과 늪지에서 걷기에 적합하다. 무스는 긴 다리를 가지고 있어서 비교적 깊은 눈이나 습지를 건널 수 있다. 이들 초식 동물들은 바깥쪽으로 벌어진 발굽을 가지고 있어서 무게를 분산시킬 수 있기 때문에 습지를 쉽게 헤쳐 나갈 수 있다<15>. 그들의 장점과 상관없이, 시베리아 겨울은 여전히 이 포유류들에게 고통스럽다: “북부 유제류(역주: 발굽이 있는 동물)는 겨울 동안 체중과 몸 상태가 서서히 감소한다<16>.” 매머드들과 많은 다른 종류의 포유동물들은 시베리아의 여름 환경에 적합하지 않다. 시베리아에서 수백 만 마리의 매머드들은 무엇을 하고 있었을까?
Starving to death amidst abundance(풍요 가운데 굶어 죽다)
비록 빙하 시대의 포유동물들이 추운 겨울과 늪지대가 있는 여름 동안 살아남았을지라도, 그들은 여전히 식량을 찾아야 했을 것이다. 시베리아의 여름에는 풍부한 식물이 있었다. 몇몇 과학자들은 매머드와 다른 동물들이 이런 풍성한 식물들로 살아남을 수 있었다고 믿는다. Farrand<17>는 다음과 같이 낙관적으로 얘기하고 있다:
고위도의 북극 지역에 가본 적이 없는 사람들은 초목, 꽃, 관목 및 난쟁이나무 등 비교적 울창한 초목에 대한 개념을 거의 가지고 있지 않은 것으로 보입니다. 그리고 하루 24시간 계속해서 햇빛이 비치는 놀라운 광경 역시 상상하지 못하겠죠!”
Farrand의 논리에는 한가지 심각한 문제가 있다. 실제 수 백만 마리의 포유류들이 풀과 작은 관목들을 먹었지만 오늘날의 시베리아 초목은 주로 늪지와 소택지 식물들이다. 현재의 식물들에는 이 거대한 동물들의 건강에 필요한 영양분이 부족하다<18>. 현재 툰드라의 남쪽에 있는 타이가(taiga) 숲 식물들 또한 이러한 동물들을 위한 영양분을 공급하기에 충분하지 않다<19>. 타이가 식물을 포함하여 오늘날 시베리아에서 자라는 가문비나무, 소형의 오리나무, 자작나무 등의 나무 껍질에는 포유류에게 유독한 독소가 많이 들어 있다<20>. 동물들은 이러한 빈약한 영양 공급 속에서 생존 할 수 없다. 과학자들은 이러한 식량 부족을 “생산성 역설”이라고 명명했다<21>. 다시 말해, 오늘날 시베리아에는 매머드와 다른 동물들을 위한 식량은 거의 없다! 작은 초원 지대가 있는 것은 사실이지만, 이곳에 사는 모든 동물들에게, 특히 몸집이 큰 포유동물들을 위한 식량으로는 충분하지 않다. 털북숭이 매머드와 그들의 동료들은 외관상 풍요로운 식물이 있는 가운데 굶어 죽었을 것이다.
매머드의 식사 및 식수의 필요량을 이해한다면 문제는 더 복잡해진다. 털북숭이 매머드는 그들과 크기가 매우 비슷한 현대의 코끼리와 비교될 수 있다. 거대한 털북숭이 매머드는 하루에 400에서 650파운드(180에서 300kg)의 즙이 많은 음식을 필요로 했을 것이다<22>. 시베리아에서 이러한 식량은 어디서 나올까?
또한 털북숭이 매머드는 매일 35에서 50갤런(140에서 200리터)의 물을 마셔야 했을 것이다<23>. 겨울 동안에 시베리아의 거의 모든 물은 얼어 있다. 어떤 사람들은 이 매머드가 절벽에서 고드름을 깨기 위해 어금니를 사용하거나, 물을 얻기 위해 눈을 먹었을 수도 있다고 제안했다<24>. 그러나 눈이나 얼음을 먹는 것은 물을 얻기 위한 효율적인 방법이 아니다. 그것은 몸을 춥게 만든다. 만약 그랬다면 겨울 동안 매머드에 의해 소모된 에너지의 절반은 그들의 위장 속의 눈을 녹이고 위장을 따뜻하게 하는데 사용되었을 것이다<25>. 또한 시베리아의 평원에는 고드름 달린 절벽이 많지 않다.
The climate enigma(기후 수수께끼)
이 시점에서 분명한 것은 털북숭이 매머드가 살았을 당시 시베리아의 기후와 환경은 지금보다 훨씬 더 따뜻했다는 것이다. 그런데 역설적으로 그들은 빙하기에 살았다. 빙하기 기후에 대한 컴퓨터 모델은 질적으로 다양하지만 결과는 일관성이 있다: 즉, 시베리아는 지금보다 훨씬 더 추웠어야 했다. Arkhipov<26>는 이렇게 결론을 내린다: “빙하기와 아빙기에 시베리아의 기후는 현재보다 훨씬 추웠을 것이다.” 아빙기는 빙하기의 가장 추운 부분이다. Manabe와 Broccoli<27>는 그들의 빙하기 기후 모델을 사용하여 현재의 시베리아 겨울보다 화씨로 약 20도(섭씨 12도) 더 추운 온도를 계산했다.
많은 빙하기 컴퓨터 시뮬레이션은 심지어 시베리아에 빙상을 만들기도 한다. 좋은 예는 Dong과 Valdes의 시뮬레이션이다<28>. 더 흥미로운 일반적인 순환 모델들 중 하나는 맨 처음부터 빙상을 만드는 것을 시도한 것이다. 지구 일조량을 6%까지 줄임으로써 빙상의 성장을 유도했다. 역설인 것은 이 시뮬레이션 결과 알래스카, 시베리아의 일부 지역 및 서부 캐나다에 영구적인 적설을 만들었지만, 실제로 빙상이 형성된 지역에는 눈이 거의 덮이지 않았다. 빙하 시대 동안에 얼음이 형성된 지역과 정반대가 된 것이다:
“우리는 시뮬레이션을 통해 빙하형성을 만들었지만, 최근의 빙하 시대에 실제 빙하가 존재했던 지역 외곽에 형성 되었다<29>.”
이러한 컴퓨터 시뮬레이션들은 시베리아에 빙하가 얼마나 쉽게 형성 될 수 있는지 보여 주지만, 어떤 점에서는 매우 잘못된 것이다. 시베리아가 빙하로 뒤덮였을 때 털북숭이 매머드가 어떻게 시베리아에서 살아남을 수 있었을까?
빙하 암설은 시베리아, 알래스카 및 유콘의 산들만이 실제로 빙하로 덮여 있었음을 나타낸다. 매머드 뼈가 발견 된 저지대는 결코 빙하가 없었다! 그것은 빙하 시대에 동물들이 이 지역에서 어떻게 살 수 있었는지를 설명해 준다. 그런데 왜 이 저지대는 빙하가 되지 않았을까?
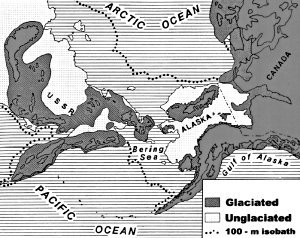
그림 2.3. 빙하기 동안 시베리아, 알래스카 및 유콘의 빙하 및 비빙하 영역. 얕은 대륙붕은 330 피트(100m)의 등심선으로 표시했다.
자기들의 모델을 정당화하기를 열망하는 일부 동일과정설 과학자들은 더 추운 빙하기 기후가 털북숭이 매머드들이 시베리아에서 살 수 있도록 도왔을 것이라고 한다. 더 추운 기온으로 인해 영구 동토층이 덜 녹게 될 것이고, 이로 인해 툰드라와 타이가 지역이 여름에 습지가 훨씬 적게 형성 되었을 것이라고 한다. 만약에 그곳이 얼은 채로 유지되었다면 이 상황은 확실히 매머드가 늪지대를 헤쳐 나가기 쉽게 했을 것이다. 그러나 더 추운 기후로 인해 생장 기간이 짧아져서 식량과 물이 많이 부족한 상태가 된다. 게다가 일부 동물들은 추운 빙하 시대보다 훨씬 덜 추운 오늘날의 추위에서도 견디기 힘들어 한다.
다시 한번, 모든 동물들이 그렇게 추운 빙하 시대에 무엇을 먹었을까? 그러한 기후에서는 식용 식물이 많이 자라지 않는다. 더 추운 빙하기의 기온은 생산성 역설을 악화시킨다. Charles Schweger<30>는 이 문제를 이렇게 설명한다:
아마도 일차 생산성이 여전히 낮았던 시기에 북쪽의 빙하 가장자리에[빙하 근처에] 더 크고 더 다양한 유제류[발굽 가진 포유동물]가 과거에 존재했다는 것은 역설적이다. 지나치게 단순화해서 질문을 한다면, 빙하기 때 Beringia 지역의 겉보기에 불가능한 조건 하에서 어떻게 맘모스가 살아 갈 수 있었을까? [Beringia는 동 시베리아, 알래스카 및 유콘 지역이다.]
그 곳에 살았던 동물들을 관찰할 때 기후의 역설은 더 악화된다. 물론, 죽은 뼈는 말할 수 없지만, 그들은 그들이 살았던 환경에 대한 단서를 제공할 수 있다. 포유류의 다양성과 그들의 생태적 요구 사항은 오늘날과는 환경이 매우 차이 나며 빙하기 기후 시뮬레이션과도 많이 차이가 난다. 빙하기 동안의 시베리아 환경은 동 아프리카 세렝게티의 환경과 비교할 만 하다!<31>
실제적으로 이 모든 거대한 포유류는 다양한 초본 식물, 즉 주로 목초를 먹는 초식 동물이었다는 사실에 기초하여 우리는 시베리아의 환경을 재현 할 수 있다. 이에 대한 추론의 기초는 다음과 같다. (1) 생존한 포유 동물의 먹이 선호도. (2) 매머드는 코끼리와 유사하다. (3) 냉동 사체의 위 내용물에 풍부한 풀을 함유하고 있다. (4) 미국 남서부의 동굴에 매머드의 배설물이 보존되어 있다<32>. 이것으로부터 우리는 시베리아가 빙하기 동안에 거대한 목초지였다고 결론을 내릴 수 있다! 내가 털북숭이 매머드 시대의 시베리아의 환경을 가장 잘 이해하고 있다고 생각하는 Dale Guthrie는 시베리아를 매머드 스텝지대라고 부른다. 스텝지대란 풀이 무성하고 나무가 거의 없는 반 건조 초원이다. 오늘날 세계에는 두 개의 주요한 스텝지대가 있다: (1) 미국 로키 산맥의 동쪽에 있는 고원 지대. (2) 중앙 아시아 산맥의 북쪽 경사면.
몇몇 작은 포유류들은 Guthrie의 매머드 스텝지대 이론을 특히 더 강화시킨다. 발견된 작은 포유류의 뼈로는 오소리와 흰족제비가 있다. 그들은 중앙 북아메리카의 록키 산맥 동쪽의 고원 평야와 같은 짧은 풀밭 평원에 살고 있다<33>. 이 동물들은 영구 동토층이 거의 없는 꽤 깊은 곳에 굴을 파고 살아간다. 초지 환경에는 북극 땅 다람쥐가 넓게 분포하고 있음이 나타났는데, 이 북극 땅 다람쥐는 개방된 지역에 살며, 에너지가 풍부한 씨앗과 단백질이 풍부한 초본을 필요로 한다<34>.
이 큰 동물 집단이 계속 유지되려면 초원에는 다른 초원과 비슷한 다양한 식물이 풍부하게 있어야 한다. Guthrie<35>는 식물 역시 매우 다양했다는 논리적인 결론을 내렸다. 그 다양성은 오늘날의 식물에 없는 다양성이다<36>. 키 큰 관목과 나무들의 작은 군락들이 이 거대한 초원 지대를 가능하게 했다. 이것들은 영구동토층에 보존 되어 있다<37>. 수많은 사람들의 의견을 바탕으로, Guthrie는 시베리아, 알래스카 및 유콘 뿐만 아니라 유럽과 러시아 서부가 하나의 거대한 초원이라고 결론을 내린다. 매머드 스텝지대는 북반구의 비 빙하 지역의 대부분을 덮은 것으로 추정된다.
그러한 다양한 식물이 있는 초원이 존재하기 위해서는 토양이 매우 비옥했음이 틀림없다<38>. 시베리아 포유동물의 뼈에는 피카(pica)라고 불리는 씹은 표시(chew marks)가 없는 것을 볼 때, 거기에 풍부한 양분을 가진 비옥한 토양이 존재했다고 추정할 수 있다<39>. 오늘날 많은 환경에서 흙이 충분한 영양분을 함유하고 있지 않기 때문에 작은 동물들은 그들의 음식에서 충분한 미네랄을 얻지 못하고 있다. 그래서 그들은 부족한 영양분을 얻기 위해 죽은 동물의 뼈를 씹는다.
포유동물의 성장 패턴 또한 비옥한 초지를 암시한다<40>. 당시의 대부분의 포유류들은 잘 차려 입은 거인들로 묘사되어 왔다. 말, 털북숭이 코뿔소와 털북숭이 매머드는 먼 남쪽의 동물보다 작았지만, 다른 모든 동물은 오늘날의 동물보다 훨씬 컸다. 게다가, 많은 동물이 가진 덥수룩한 목 털, 무거운 뿔, 긴 어금니 및 거대한 뿔로 볼 때 경쟁이 거의 없고 성장기가 길고 양질의 서식지가 있었던 것으로 추정된다<41>. 토양 영양분은 특히 동물의 크기와 사회 공동체의 크기에 영향을 끼친다<42>. Bergmann의 법칙은 기후가 더 추울수록 동물이 더 크다고 말하지만, 다른 사람들은 Bergmann의 법칙에 강력히 반대하며, 동물의 크기는 성장기 동안의 먹이의 질에 비례한다고 주장한다<43>. Guthrie<44>는 알래스카의 돌시프(북미 북서부 산악 지방산(產)인 털이 횐 야생 양)에 대한 실험을 통해 동물의 크기와 먹이와의 연관성을 보여주었다: 먹이가 많을수록 더 크게 자랐다.
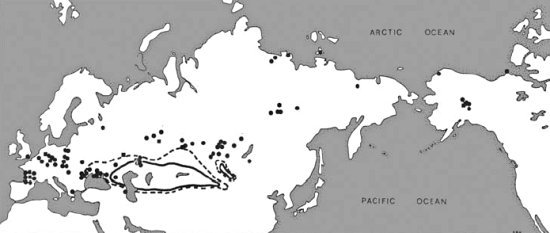
그림 2.4. 사이가산양(saiga antelope)의 분포: 현재(실선), 역사적(점선), 그리고 빙하기(점)
이렇게 다양한 초본 식물을 유지하려면 따뜻한 토양과 급격한 봄 성장으로 길게 자라는 계절이 필요하다<45>. 이러한 기후는 6월 중순부터 7월 초순까지 북부 시베리아에서 녹색새싹 조차 찾을 수 없는 현재의 환경과는 크게 다르다<46>. 스텝기후는 초원 유지에 필요한 기후의 일종인 비가 많은 봄, 건조한 늦여름과 초가을을 가지고 있다. 결과적으로, 매머드 시대에 시베리아에도 이와 비슷한 강수량 패턴을 예상하는 것이 합리적이다.
초지 생물 자원의 90%가 지표 아래에 뿌리를 두고 있다. 식물은 눈이 녹고 토양이 따뜻해질 때까지 신선한 싹을 발아시킬 수 없다. 스텝 환경은 토양 수분 함량이 적고 겨울 강설량이 적은 특징을 갖고 있다<47>. 이것은 또한 겨울 강설량이 적었고 일찍 녹았음을 의미하며, 온화하고 비교적 건조한 겨울은 봄을 재촉한다.
사이가 영양, 큰 뿔 양, 돌시프, 늑대와 같이 많은 눈을 견딜 수 없는 여러 동물의 존재는 겨울 강설량이 적었음을 추정하게 한다<48>. 들소는 머리와 뿔로 눈을 긁어내는데 아주 효율적이지만, 눈이 2피트를 넘으면 음식을 충분히 얻는데 어려움을 겪는다. 영구 동토층에서 많은 들소 유해들이 발견되는 중앙 알래스카 지역에는 눈이 보통 3피트 이상 쌓인다. 들소는 바람이 부는 작은 지역을 제외하고는 알래스카 중부에 살 수 없다<49>.
요약하면, 시베리아 동물의 생태는 비옥한 토양을 가진 훨씬 더 다양한 식물이 있었음을 추정하게 한다. 또한 이것은 적은 강설량과 긴 성장 시기를 가진 비교적 온화한 겨울을 의미한다. 이러한 조건은 현대 환경과 기후와 뚜렷하게 다르며, 이것은 동일과정설에 기반을 둔 빙하기 기후에 대한 컴퓨터 시뮬레이션으로는 설명할 수 없다.
참고문헌
1. Stuart, A.J., Mammalian extinctions in the Late Pleistocene of northern Eurasia and North America, Review of Biology 66:508, 1991.
2. Knystautas, A., The natural history of the USSR, McGraw-Hill, New York, p. 27, 1987.
3. Vereshchagin, N.K., and G.F. Baryshnikov, Quaternary mammalian extinctions in Northern Eurasia; in: Quaternary extinctions: A prehistoric revolution, P.S. Martin and R.G. Klein (Eds.), University of Arizona Press, Tucson, AZ, p. 492, 1984.
4. Digby, B., The mammoth and mammoth-hunting in north-east Siberia, H.F. & G. Witherby, London, p. 195, 1926.
5. Soffer, O., The upper paleolithic of the Central Russian Plain, Academic Press, New York, 1985.
6. Guthrie, R.D., Frozen fauna of the mammoth steppe — The story of Blue Babe, University of Chicago Press, Chicago, IL, p. 246, 1990.
7. Howorth, H.H., The Mammoth and the flood — An attempt to confront the theory of uniformity with the facts of recent geology, Sampson Low, Marston, Searle, & Rivington, London, reproduced by The Sourcebook Project, Glen Arm, Maryland, p. 62–63, 1887.
8. Haynes, G., Mammoths, mastodonts, and elephants, Cambridge University Press, Cambridge, NY, p. 97, 1991.
9. Haynes, G., The mountains that fell down: Life and death of heartland mammoths; in: Megafauna and man — Discovery of America’s heartland, L. Agenbroad, J.I. Mead, and L.W. Nelson (Eds.), The Mammoth Site of Hot Springs, South Dakota, Inc., Hot Springs, SD, scientific papers, vol. 1, p. 25–26, 1990.
10. Digby, Mammoth and mammoth-hunting, p. 15–16. Vereshchagin and Baryshnikov, Quaternary mammalian extinctions, p. 492.
11. Pfizenmayer, E.W., Siberian man and mammoth, Blackie & Sons, London, 1939.
12. Tolmachoff, I.P., The carcasses of the mammoth and rhinoceros found in the frozen ground of Siberia, Transactions of the American Philosophical Society 23:57, 1929.
13. Farrand, W.R., Frozen mammoths and modern geology, Science 133:734, 1961.
14. Vereshchagin, N.K., The mammoth “cemeteries” of north-east Siberia, Polar Record 17(106):12, 1974.
15. Guthrie, R.D., Mammals of the mammoth steppe as paleoenvironmental indicators; in: Paleoecology of Beringia, D.M. Hopkins, J.V. Matthews Jr., C.E. Schweger, and S.B. Young (Eds.), Academic Press, New York, p. 310, 1982.
16. Ibid., p. 320.
17. Farrand, W.R., Frozen mammoths, Science 137:451, 1962.
18. Chapin, III, F.S., G.R. Shaver, A.E. Giblin, K.J. Nadelhoffer, and J.A. Laundre, Responses of Arctic tundra to experimental and observed changes in climate, Ecology 76(3):694, 1995.
19. Sher, A.V., Late-Quaternary extinction of large mammals in northern Eurasia: A new look at the Siberian contribution; in: Past and future rapid environmental changes: The spatial and evolutionary responses of terrestrial biota, B. Huntley, W. Cramer, A.V. Morgan, H.C. Prentice, and J.R.M. Allen (Eds.), Springer, New York, p. 322, 1997.
20. Guthrie, R.D., and M.L. Guthrie, On the mammoth’s dusty trail, Natural History 99(7):40, 1990.
21. Hopkins, D.M., J.V. Matthews Jr., C.E. Schweger, and S.B. Young (Eds.), Paleoecology of Beringia, Academic Press, New York, 1982.
22. Vereshchagin, N.K., and G.F. Baryshnikov, Paleoecology of the mammoth fauna in the Eurasian Arctic; in: Paleoecology of Beringia, D.M. Hopkins, J.V. Matthews Jr., C.E. Schweger, and S.B. Young (Eds.), Academic Press, New York, p. 269, 1982.
23. Webb, S.D., A brief history of new world Proboscidea with emphasis on their adaptations and interactions with man; in: Proboscidean and paleoindian interactions, J.W. Fox, C.B. Smith, and K.T. Wilkins (Eds.), Baylor University Press, Waco, TX, p. 27, 1992.
24. Digby, Mammoth and mammoth-hunting, p. 52. Vereshchagin and Baryshnikov, Quaternary mammalian extinctions, p. 490.
25. Olivier, R.C.D., Ecology and behavior of living elephants: Bases for assumptions concerning the extinct woolly mammoths; in: Paleoecology of Beringia, D.M. Hopkins, J.V. Matthews Jr., C.E. Schweger, and S.B. Young (Eds.), Academic Press, New York, p. 303, 1982.
26. Arkhipov, S.A., Environment and climate of Sartan maximum and late glacial in Siberia; in: Late glacial and postglacial environmental changes — Quaternary, Carboniferous-Permian, and Proterozoic, I.P. Martini (Ed.), Oxford University Press, New York, p. 54, 1997.
27. Manabe, S., and A.J. Broccoli, The influence of continental ice sheets on the climate of an ice age, Journal of Geophysical Research 90(C2):2167–2190, 1985.
28. Dong, B., and P.J. Valdes, Sensitivity studies of Northern Hemisphere glaciation using an atmospheric general circulation model, Journal of Climate 8:2471–2496, 1995.
29. Phillips, P.J., and I.M. Held, The response to orbital perturbations in an atmospheric model coupled to a slab ocean, Journal of Climate 7:780, 1994.
30. Schweger, C.E., Primary production and the Pleistocene ungulates — The productivity paradox; in: Paleoecology of Beringia, D.M. Hopkins, J.V. Matthews Jr., C.E. Schweger, and S.B. Young (Eds.), Academic Press, New York, p. 219, 1982.
31. Guthrie, Mammals, p. 313. Bocherens, H., G. Pacaud, P.A. Lazarev, and A. Mariotti, Stable isotope abundances (13C, 15N) in collagen and soft tissues from Pleistocene mammals from Yakutia: Implications for the palaeobiology of the mammoth steppe, Palaeogeography, Palaeoclimatology, Palaeoecology 126:31, 1996.
32. Guthrie, Mammals, p. 307–326. Guthrie, Frozen fauna. Haynes, Mammoths, mastodonts, and elephants, p. 59.
33. Guthrie, Frozen fauna, p. 248–249. Pielou, E.C., After the Ice Age — The return of life to glaciated North America, University of Chicago Press, Chicago, IL, p. 151, 1991.
34. Guthrie, Mammals, p. 311.
35. Guthrie, Mammals, p. 315.
36. Wright Jr., H.E., and C.W. Barnosky, Introduction to the English edition; in: Late Quaternary environments of the Soviet Union, A.A. Velichko (Ed.), University of Minnesota Press, Minneapolis, MN, p. xiii–xxii, 1984. Guthrie, R.D., Mosaics, allelochemics and nutrients — An ecological theory of late Pleistocene megafaunal extinctions; in: Quaternary extinctions: A prehistoric revolution, P.S. Martin and R.G. Klein (Eds.), University of Arizona Press, Tuscon, AZ, p. 259–297, 1984.
37. Tolmachoff, Carcasses, p. 47. Kaplina, T.N., and A.V. Lozhkin, Age and history of accumulation of the “ice complex” of the maritime lowlands of Yakutia; in: Late Quaternary environments of the Soviet Union, A.A. Velichko (Ed.), University of Minnesota Press, Minneapolis, MN, p. 147, 1984. Anderson, P.M., Late Quaternary vegetational change in the Kotzebue Sound area, Northwestern Alaska, Quaternary Research 24:307–321, 1985. Sher, Late-Quaternary extinction, p. 322.
38. Guthrie, Mosaics, allelochemics and nutrients, p. 267.
39. Guthrie, Frozen fauna, p. 215–219.
40. Guthrie, Mammals, p. 307–326.
41. Guthrie, Mammals, p. 309.
42. Guthrie, Mosaics, allelochemics and nutrients.
43. Geist, V., Bergmann’s rule is invalid, Canadian Journal of Zoology 65:1035–1038, 1987. Guthrie, Mosaics, allelochemics and nutrients, p. 269, 271.
44. Guthrie, Mosaics, allelochemics and nutrients, p. 271.
45. Guthrie, Mammals, p. 322–324.
46. Howorth, H.H., The mammoths in Siberia, Geological Magazine 7:553, 1880.
47. Berman, D., S. Armbruster, A. Alfimov, and M. Edwards, Subarctic steppe communities in Beringia; in: Bridges of the science between North America and the Russian Far East, 45th Arctic science conference, section 2 — Beringia revisited: Recent discoveries and interpretations, p. 10, 1994.
48. Guthrie, Frozen fauna, p. 201.
49. Guthrie, Frozen fauna.
 Frozen in Time(제3장) 빙하기의 신비 - Michael J. Oard 글
Frozen in Time(제3장) 빙하기의 신비 - Michael J. Oard 글
 Frozen in Time(제1장) 시베리아에 냉동된 매머드 사체 - Michael...
Frozen in Time(제1장) 시베리아에 냉동된 매머드 사체 - Michael...